

- مقاله
- دسترسی آزاد
- منتشر شده:
تنوع مورفولوژیکی گرده و هاگ در یک موزاییک جنگل-کشاورزی کوهستانی تحت تأثیر انسان در شمال تایلند
گزارشهای علمی حجم ۱۶ ، شماره مقاله: ۶۷۹۴ ( ۲۰۲۶ )
چکیده
ریختشناسی گرده و هاگ، دادههای مرجع طبقهبندی ضروری را برای مطالعات فلوریستیک و زیستمحیطی در مناطق گرمسیری، جایی که مجموعه دادههای مقایسهای مدرن محدود هستند، فراهم میکند. این مطالعه ویژگیهای ریختشناسی گرده و هاگهای بازیابی شده از یک پروفیل خاک کمعمق در یک جنگل برگریز مخلوط تخریبشده در پارک ملی سری نان، شمال تایلند را مستند میکند. با استفاده از یک پروتکل استخراج غیر استولیز و نمونهبرداری سیستماتیک از یک پروفیل ۳۰ سانتیمتری، گرده و هاگهای نماینده ۳۷ خانواده گیاهی، از جمله لیکوفیتها، بریوفیتها، مونیلوفیتها، بازدانگان و نهاندانگان شناسایی شدند. گونههای تولیدکننده هاگ، بهویژه مونیلوفیتها، بر این مجموعه تسلط دارند، در حالی که گرده نهاندانگان شامل عناصر درختی و غیر درختی است. بیش از ۱۰۰ مورفوتیپ بر اساس نوع روزنه، تزئینات اگزین، اندازه و تقارن، که توسط میکروسکوپهای نوری با وضوح بالا و توصیفات استاندارد مورفوتیپ پشتیبانی میشوند، توصیف شدهاند. مجموعه دادههای حاصل، چارچوب مرجع پالینولوژیکی منطقهای را برای شمال تایلند و مناطق گرمسیری جنوب شرقی آسیا گسترش میدهد و از شناسایی طبقهبندی منسجم در مطالعات پالینولوژیکی، فلوریستیک و پالئواکولوژیکی مقایسهای، به ویژه در موزاییکهای جنگل-کشاورزی تحت تأثیر انسان، پشتیبانی میکند.
محتوای مشابه توسط دیگران مشاهده میشود

مقدمه
ریختشناسی گرده و هاگ همچنان برای شناسایی طبقهبندی در پالینولوژی، بهویژه در مناطق گرمسیری که تنوع فلوریستیک بالا است و مجموعههای مرجع اغلب ناقص هستند، اساسی است ۱ ، ۲ ، ۳ ، ۴٫ در حالی که اطلسهای جامع ریختشناسی گرده و هاگ برای بسیاری از مناطق گرمسیری در حال حاضر در دسترس هستند (به عنوان مثال ۱ ، ۳ ، ۵ )، مجموعه دادههای کالیبره شده منطقهای برای اطمینان از شناسایی طبقهبندی سازگار در محیطهای رسوبی مختلف، پروتکلهای آزمایشگاهی و زمینههای اکولوژیکی ضروری هستند. در مناظر تحت تأثیر تغییر کاربری زمین و تخریب اکولوژیکی – که در آن موجودیهای گیاهشناسی ممکن است محدود باشند – ریختشناسی پالینومورف، بایگانی پایداری از ترکیب پوشش گیاهی و یک خط مستقل از شواهد برای ارزیابی اکوسیستم فراهم میکند ۶ ، ۷ ، ۸٫ مجموعه دادههای ریختشناسی کالیبره شده منطقهای همچنین برای کارهای میان رشتهای در بومشناسی مرمت، باستانشناسی و مطالعات مرتبط با آب و هوا، که در آن اطمینان در سطح تاکسون میتواند استنتاج پاییندست را تغییر دهد ۹ ، ۱۰ ، به طور فزایندهای اهمیت دارند .
مناظر کوهستانی شمال تایلند شامل جنگلهای برگریز مخلوط است که یک گذار اکوتونال بین جنگلهای همیشه سبز و جنگلهای خشک فصلی دیپتروکارپ را تشکیل میدهند. این سیستمها از عناصر فلوریستیک مرتبط با مناطق هند و برمه و چین و هیمالیا پشتیبانی میکنند [۱۱ ، ۱۲ ، ۱۳] ، با این حال، آنها دستخوش تبدیل و تکهتکه شدن گستردهای شدهاند، به ویژه با گسترش کشاورزی مانند کشت ذرت و تککشتی پارا کائوچو [۱۴ ، ۱۵] . چنین تغییر کاربری زمین میتواند تولید، انتقال و رسوب گرده را تغییر دهد و باران گرده محلی و مجموعههای پالینومورف حفظ شده در خاکها و رسوبات را اصلاح کند [۱۶] . در عین حال، کاربرد عملی سوابق پالینومورف به شدت به در دسترس بودن منابع مورفولوژی منطقهای مستند بستگی دارد که امکان شناسایی مداوم را در مطالعات و تحلیلگران فراهم میکند.
علیرغم در دسترس بودن اطلسهای گردهشناسی متمرکز بر ریختشناسی و مطالعات گردهشناسی مدرن از جنوب شرقی آسیا و مناطق گرمسیری مجاور (مثلاً ۱ ، ۳ ، ۵ )، اکثر منابع موجود بر جنگلهای نسبتاً دستنخورده، پیتزارها یا باران گرده سطحی تأکید دارند. در مقابل، مستندسازی متمرکز بر ریختشناسی از موزاییکهای جنگلی-کشاورزیِ تحت تأثیر فعال انسان در شمال تایلند همچنان محدود است. این مطالعه با مستندسازی ریختشناسی پالینومورف از یک پروفیل رسوب ۳۰ سانتیمتری جمعآوریشده در پارک ملی سری نان، استان نان، به این شکاف میپردازد. با استفاده از یک رویکرد استخراج غیر استولیز و نمونهبرداری با وضوح بالا در فواصل ۳ سانتیمتری، ما یک مرجع مورفوتایپ مبتنی بر تصویر ارائه میدهیم که گردههای درختی، گردههای غیر درختی و هاگها را در بر میگیرد و توصیفاتی بر اساس نوع روزنه، تزئینات اگزین، اندازه و تقارن ارائه میدهد. هدف ما گسترش چارچوب مرجع پالینولوژیکی منطقهای برای آسیای جنوب شرقی گرمسیری و بهبود تکرارپذیری در شناسایی طبقهبندی برای کاربردهای پالینولوژیکی و محیطی آینده است.
یک بازسازی دیرینهاکولوژیکی گستردهتر بر اساس توالی رسوب طولانیتر از منطقه مورد مطالعه، شامل مدلسازی سن-عمق و تفسیر اختلال کاربری زمین در طول زمان، قبلاً منتشر شده است [۱۷] . مطالعه حاضر بر مستندسازی مورفولوژی محور (مورفوتایپها، ویژگیهای تشخیصی و عکسهای میکروسکوپی با وضوح بالا) مشتق شده از یک پروفیل ۳۰ سانتیمتری با استفاده از یک گردش کار غیر استولیز تمرکز دارد و یک منبع مرجع قابل استفاده مجدد برای پشتیبانی از شناسایی شفاف و تسهیل کاربردهای پاییندست، از جمله فلوریستیک مقایسهای و توسعه روششناختی، ارائه میدهد. بنابراین، تفسیرها به مستندسازی مورفولوژیکی و ارتباطات زمینهای محدود میشوند و بازسازی مستقلی از تاریخ پوشش گیاهی محسوب نمیشوند.
روششناسی
منطقه مورد مطالعه
پارک ملی سری نان، واقع در استان نان در شمال تایلند، تقریباً ۱۰۲۴۳۸ هکتار را در مناطق نا موئن، نا نوی و ویانگ سا در بر میگیرد (شکل ۱ ). این پارک دارای یک زمین کوهستانی ناهموار است که با پشتههای متناوب و درههای بین کوهی مشخص میشود و بالاترین ارتفاع آن به ۱۲۳۴ متر از سطح دریا میرسد. این پیچیدگی توپوگرافی از تنوع زیستی بالا و اکوسیستمهای جنگلی متنوع پشتیبانی میکند و در عین حال به عنوان یک حوزه آبخیز حیاتی برای رودخانه نان، یکی از شاخههای اصلی رودخانه چائو فرایا، عمل میکند.
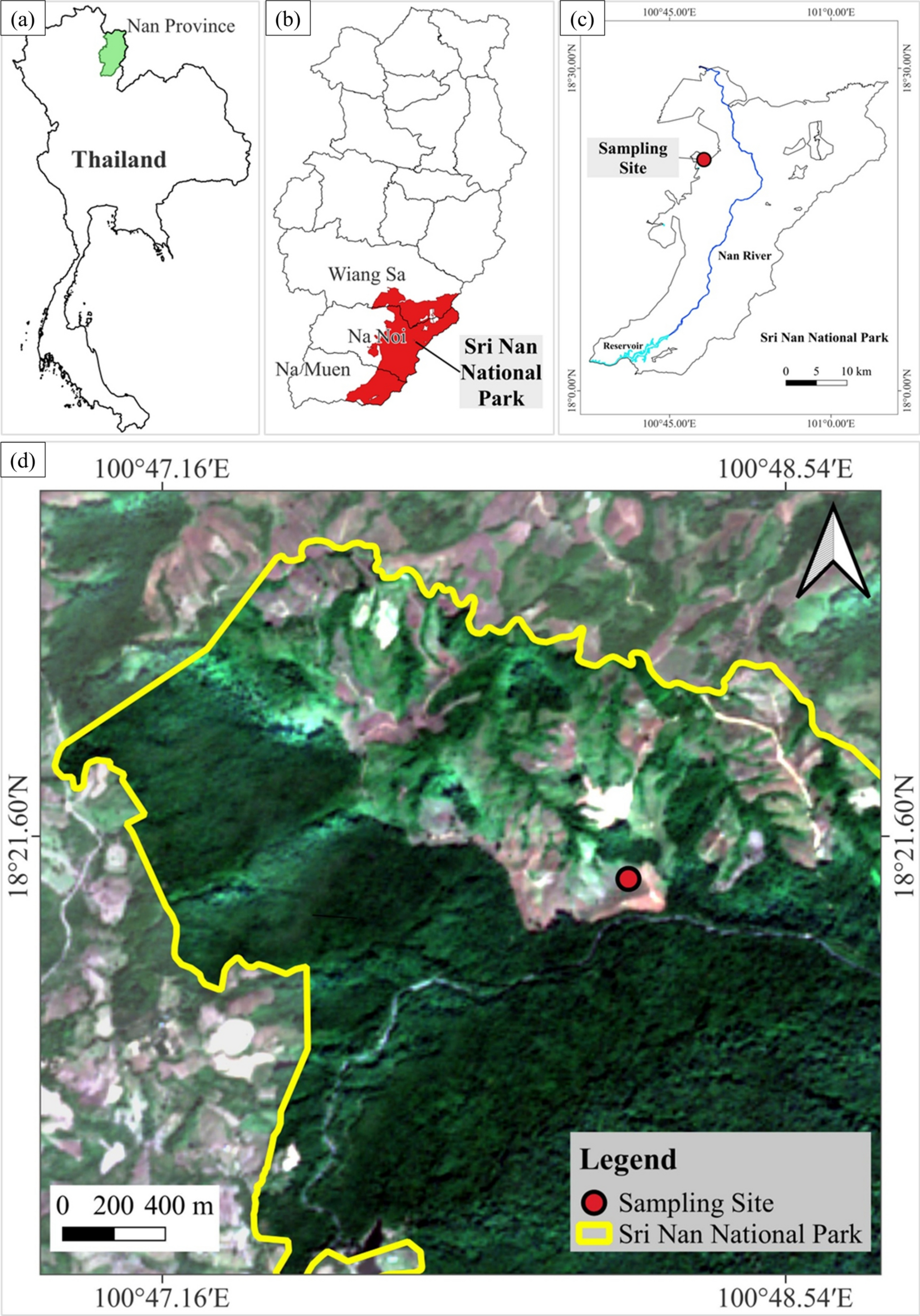
موقعیت جغرافیایی محل نمونهبرداری گردهشناسی در پارک ملی سری نان، شمال تایلند. ( الف ) زمینه ملی که موقعیت تایلند را نشان میدهد؛ ( ب ) استان نان که سه منطقه تحت پوشش منطقه مورد مطالعه را برجسته میکند: ویانگ سا، نا نوی و نا موئن؛ ( ج ) مرز پارک؛ ( د ) تصاویر ماهوارهای که منطقه نمونهبرداری را نشان میدهد. دایره قرمز محل دقیق نمونهبرداری را نشان میدهد که در نزدیکی اکوتون بین جنگل نسبتاً دستنخورده و زمینهای دستخورده انسانی واقع شده است. همه نقشهها با استفاده از نرمافزار QGIS 3.22.10 (QGIS.org، ۲۰۲۲؛ http://qgis.org ) ایجاد شدهاند .
آب و هوا به طور مشخص فصلی است، با یک فصل گرم (فوریه-آوریل)، یک فصل بارانی (مه-اکتبر) و یک فصل خنک (نوامبر-ژانویه) و میانگین دمای سالانه تقریباً ۲۴ درجه سانتیگراد ۱۸٫ پوشش گیاهی عمدتاً از جنگلهای همیشه سبز و برگریز تشکیل شده است ۱۹٫ جنگلهای همیشه سبز شامل انواع همیشه سبز خشک و تپهای هستند که گونههایی مانند Anisoptera costata ، Dipterocarpus alatus ، Afzelia xylocarpa و Pinus kesiya در آنها غالب هستند . جنگلهای برگریز شامل انواع مخلوط برگریز و خشک dipterocarp مانند Tectona grandis ، Xylia xylocarpa ، Shorea obtusa ، Lagerstroemia floribunda و گونههای مختلف بامبو هستند.
نقشه موقعیت مکانی با استفاده از QGIS نسخه ۳٫۲۲٫۱۰ ۲۰ تهیه شد . مرزهای اداری از طریق تبادل دادههای بشردوستانه از دفتر هماهنگی امور بشردوستانه سازمان ملل متحد ۲۱ ، بر اساس مجموعه دادههای ارائه شده توسط اداره نقشهبرداری سلطنتی تایلند، به دست آمد. شبکههای جریان طبیعی از اداره منابع آب تایلند ۲۲ به دست آمد . تصاویر ماهوارهای که در ۱۶ نوامبر ۲۰۲۴ به دست آمد، از محصولات Level-2A Sentinel-2A ۲۳ گرفته شده است .
نمونهبرداری
جنگلهای برگریز مخلوط، نوع پوشش گیاهی غالب در منطقه مورد مطالعه هستند. این مطالعه به بررسی ریختشناسی گرده از نمونههای خاک جمعآوریشده در یک منطقه تخریبشده – که قبلاً پوشیده از جنگلهای برگریز مخلوط بود – میپردازد که از آن زمان به کشت ذرت تبدیل شده است (از نوامبر ۲۰۲۴). این مکان که در ۱۸٫۳۵۸۵۴۱° شمالی، ۱۰۰٫۸۰۳۸۰۲° شرقی و ۵۷۹ متر بالاتر از سطح دریا (شکل ۱ ) واقع شده است، با استفاده از عکسهای هوایی، تصاویر ماهوارهای و دادههای کاربری زمین و پوشش زمین شناسایی شد. خاک از نوع شن لومی با رنگ قهوهای مایل به قرمز تا قهوهای است. پوشش گیاهی باقیمانده اطراف شامل گونههای Dipterocarpus ، با درختان رایجی مانند Xylia xylocarpa ، Shorea obtusa ، Pterocarpus macrocarpus و Bombax ceiba است . لایه زمین تحت سلطه Chromolaena odoratum و Pennisetum polystachyon است . هیچ بررسی پوشش گیاهی مستقل یا اعتبارسنجی کمی پوشش زمین به موازات نمونهبرداری گردهشناسی انجام نشد. توصیف پوشش گیاهی و کاربری اراضی اطراف صرفاً برای زمینهسازی وقوع مورفوتیپ ارائه شده است، نه برای امکان کالیبراسیون مستقیم نمایش گرده.
این مطالعه بر اساس یک پروفیل خاک کمعمق از یک مکان انجام شده است و هدف آن ارزیابی تغییرپذیری مکانی یا نشان دادن ناهمگونی کامل موزاییک جنگل-کشاورزی در منطقه مورد مطالعه نیست. رسوب با استفاده از یک مته دستی تا عمق ۳۰ سانتیمتر جمعآوری شد، سپس در فواصل ۳ سانتیمتری نمونهبرداری شد که منجر به ۱۱ نمونه فرعی شد. همه نمونهها مهر و موم شده و برای تجزیه و تحلیل گردهشناسی به دانشگاه ماهیدول منتقل شدند.
تجزیه و تحلیل پالینولوژیکی
تجزیه و تحلیل پالینولوژیکی در این مطالعه از یک روش استخراج غیر اسیدی به دنبال ۲۴ استفاده کرد که برای حفظ یکپارچگی دانه گرده بدون استفاده از تیمارهای شیمیایی خشن انتخاب شده بود. نمونههای رسوب ابتدا همگن و سانتریفیوژ شدند و سپس مواد هیومیک با استفاده از ۱۰٪ هیدروکسید پتاسیم (KOH) با حرارت دادن و شستشوی مکرر حذف شدند. جداسازی چگالی با استفاده از کلرید روی (ZnCl2) با وزن مخصوص ۱٫۹ گرم بر سانتیمتر مکعب برای جداسازی بخش گرده انجام شد . در موارد تجمع مواد آلی، ترکیبی از KOH و متافسفات سدیم تحت حرارت برای تجزیه ماتریکس استفاده شد. باقیماندههای نهایی برای تهیه اسلاید در ۸۰٪ گلیسیرین شستشو داده شده و به حالت تعلیق درآمدند.
بررسی میکروسکوپی با بزرگنمایی ۴۰۰ برابر با استفاده از میکروسکوپ نوری عبوری انجام شد. دانههای گرده بر اساس ویژگیهای ریختشناسی با مراجعه به منابع معتبر (مثلاً ۳ ، ۵ ، ۲۵ ، ۲۶ ) و پایگاههای داده آنلاین از جمله PalDat و اطلس گرده و هاگ استرالیا شناسایی شدند. شناسایی طبقهبندی تا سطح جنس انجام شد، جایی که ویژگیهای ریختشناسی تشخیصی اجازه میدادند. هنگامی که شناسایی در سطح جنس امکانپذیر نبود، مورفوتایپها به سطح خانواده اختصاص داده شده و به عنوان gen. indet ثبت شدند. همه انواع گرده بیشتر به عنوان گرده درختی (AP) یا گرده غیر درختی (NAP) طبقهبندی شدند. شمارههای مورفوتایپ (مثلاً Pteris sp.1-sp.7) در هر گروه طبقهبندی منحصر به فرد هستند و بر اساس تفاوتهای مداوم در پیکربندی روزنه، تزئینات اگزین و اندازه تعیین میشوند. معیارهای تشخیصی دقیق برای هر مورفوتایپ در مجموعه دادههای تکمیلی ارائه شده است. این مطالعه منحصراً بر گرده و هاگ تمرکز دارد. پالینومورفهای غیر گردهای از بحث بیشتر مستثنی هستند.
نتایج
بخش نتایج، فراوانیهای مشاهدهشده، فراوانیهای نسبی و توزیع چینهشناسی گرده و هاگ را گزارش میدهد. معیارهای عدم قطعیت آماری اعمال نشده است، زیرا هدف اصلی این مطالعه مستندسازی ریختشناسی و ترکیبشناسی است نه بازسازی کمی. در سراسر نتایج، اعداد مورفوتایپ منحصراً به واحدهای مبتنی بر ریختشناسی اشاره دارند، در حالی که نامهای خانواده یا جنس گیاهشناسی، قرابت طبقهبندی استنباطشده بر اساس ادبیات گردهشناسی تطبیقی را نشان میدهند. شناساییهای سطح خانواده بدون انتساب جنس به عنوان ژن مشخص شده در شکلها و تکمیلی برچسبگذاری شدهاند.
در مجموع ۳۷ خانواده گیاهی از پروفیل رسوب ۳۰ سانتیمتری جمعآوریشده در منطقه سری نان شناسایی شدند. این خانوادهها شامل گونههای تولیدکننده هاگ، بازدانگان و نهاندانگان هستند که توسط انواع گردههای درختی و غیر درختی نشان داده میشوند.
پالینومورفهای مولد هاگ توسط ۱۴ خانواده نشان داده میشوند که شامل یک خانواده لیکوفیت (Lycopodiaceae)، یک خانواده بریوفیت (Notothyladaceae) و ۱۲ خانواده مونیلوفیت (Aspleniaceae، Blechnaceae، Osmundaceae، Pteridaceae، Dryopteridaceae، Lygodiaceae، Dennstaedtiaceae، Lindsaeaceae، Ophioglossaceae، Polypodiaceae، Vittariaceae و Cyatheaceae) هستند. علاوه بر این، هاگهای سرخسهای تریلت و مونولت نامشخص ثبت شدهاند اما به خانوادههای خاصی اختصاص داده نشدهاند. پالینومورفهای مولد هاگ در مجموع ۴۳.۴۳٪ از کل مجموعه را تشکیل میدهند.
بازدانگان توسط دو خانواده Pinaceae و Cupressaceae نشان داده میشوند که ۲٫۷۵٪ از کل پالینومورفها را تشکیل میدهند.
گرده نهاندانگان در مجموع ۲۱ خانواده را شامل میشود که شامل گونههای درختی و غیر درختی میشود. خانوادههای نهاندانگان درختی شامل Ulmaceae، Cannabaceae، Betulaceae، Fagaceae، Euphorbiaceae، Lythraceae، Sapindaceae، Ebenaceae، Bignoniaceae، Malvaceae، Anacardiaceae، Rubiaceae و Arecaceae هستند که ۱۵.۵۹٪ از مجموعه را تشکیل میدهند. گرده نهاندانگان غیر درختی تحت سلطه هشت خانواده – Amaranthaceae، Poaceae، Solanaceae، Asteraceae، Araceae، Cyperaceae، Fabaceae و Liliaceae – است که در مجموع ۳۸.۲۳٪ از مجموعه را تشکیل میدهند. برخی از خانوادهها (به عنوان مثال، Malvaceae) در هر دو دسته درختی و غیر درختی وجود دارند و بنابراین فقط یک بار در کل غنای خانواده شمارش میشوند.
از آنجا که این مواد از یک پروفیل خاک کمعمق مشتق میشوند، مجموعه پالینومورفها ورودیهایی را از منابع گیاهی محلی، فرامحلی و بالقوه منطقهای ادغام میکند و گونههای منفرد به عنوان نمایندههای مستقیم پوشش گیاهی موجود تفسیر نمیشوند.
توزیع گروههای اصلی پالینومورف بر اساس عمق
نمودار گرده (شکل ۲ ) توزیع عمودی گونههای درختی، غیر درختی و هاگ را در یک پروفیل رسوبی ۳۰ سانتیمتری از یک محیط جنگلی برگریز مختلط تحت تأثیر انسان در منطقه مورد مطالعه نشان میدهد. در مجموع ۶۱ گونه پالینومورف شناسایی و در سه دسته عملکردی (گرده درختی، گرده غیر درختی و هاگ) گروهبندی شدند. الگوهای عمقی در اینجا به عنوان مشاهدات چینهشناسی در پروفیل نمونهبرداری شده گزارش شدهاند و به عنوان یک تاریخچه پوشش گیاهی با محدودیت زمانی تفسیر نمیشوند.
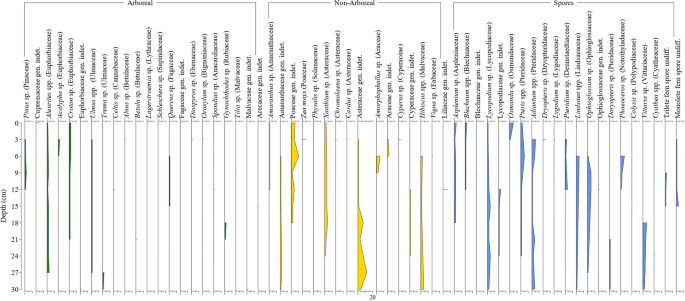
نمودار گردهشناسی نمونههای رسوب از عمق ۰ تا ۳۰ سانتیمتر در یک چشمانداز کشت جنگل-ذرت در منطقه مورد مطالعه.
صفحات فتومیکروگراف برای مورفوتایپهای مستند در شکلهای ۳ ، ۴ ، ۵ ، ۶ ، ۷ ، ۸ ، ۹ ، ۱۰ ، ۱۱ ارائه شدهاند .

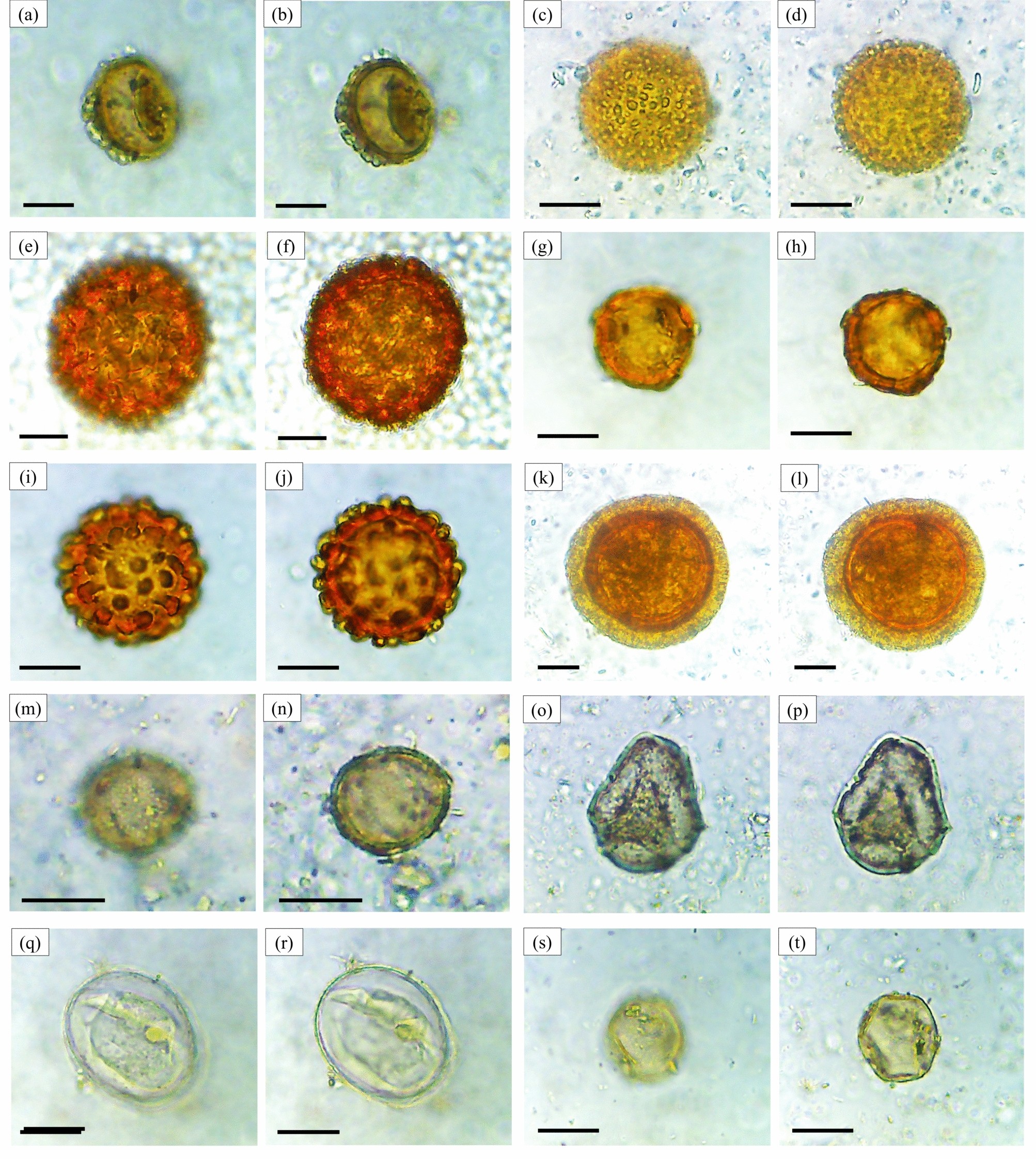
تصاویر میکروسکوپی از هاگهای لیکوفیتها، خزهها و مونیلوفیتها (بزرگنمایی ۴۰۰ ×). لیکوفیت: Lycopodium sp. ( a ، b )، Lycopodiaceae gen. indet. ( c ، d ). خزهها: Phaeoceros sp. ( e ، f ). مونیلوفیتها: Asplenium sp. ( g ، h )، Blechnum sp.1 ( i ، j )، Blechnum sp.2 ( k ، l )، Osmunda sp. ( m ، n )، Pteris sp.1 ( o ، p )، Pteris sp.2 ( q ، r )، Pteris sp.3 ( s ، t ). مقیاس = ۱۰ میکرومتر. تمایزات مورفوتیپ بر اساس تفاوت در پیکربندی روزنه، تزئینات اگزین و اندازه دانهها است (به بخش تکمیلی مراجعه کنید).

تصاویر میکروسکوپی از هاگهای مونیلوفیتها (بزرگنمایی ۴۰۰ ×). مونیلوفیتها: Pteris sp.4 ( a ، b )، Pteris sp.5 ( c ، d )، Pteris sp.6 ( e ، f )، Pteris sp.7 ( g ، h )، Adiantum sp.1 ( i ، j )، Adiantum sp.2 ( k ، l )، Adiantum sp.3 ( m ، n )، Adiantum sp.4 ( o ، p )، Doryopteris sp. ( q ، r )، Dryopteris sp. ( s ، t ). مقیاس = ۱۰ میکرومتر. تمایزات مورفوتیپ از معیارهای شرح داده شده در شکل ۳ و تکمیلی پیروی میکنند.

تصاویر میکروسکوپی از هاگها و دانههای گرده مونیلوفیتها (بزرگنمایی ۴۰۰ ×). مونیلوفیتها: Lygodium sp. ( a ، b )، Pteridium sp. ( c ، d )، Lindsaea sp.1 ( e ، f )، Lindsaea sp.2 ( g ، h )، Ophioglossum sp. ( i ، j )، Ophioglossaceae gen. indet. ( k ، l )، Colysis sp. ( m ، n )، Vittaria sp. ( o ، p )، Cyathea sp.1 ( q ، r )، Cyathea sp.2 ( s ، t ). مقیاس = ۱۰ میکرومتر. تمایزات مورفوتیپ از معیارهای شرح داده شده در شکل ۳ و تکمیلی پیروی میکنند.

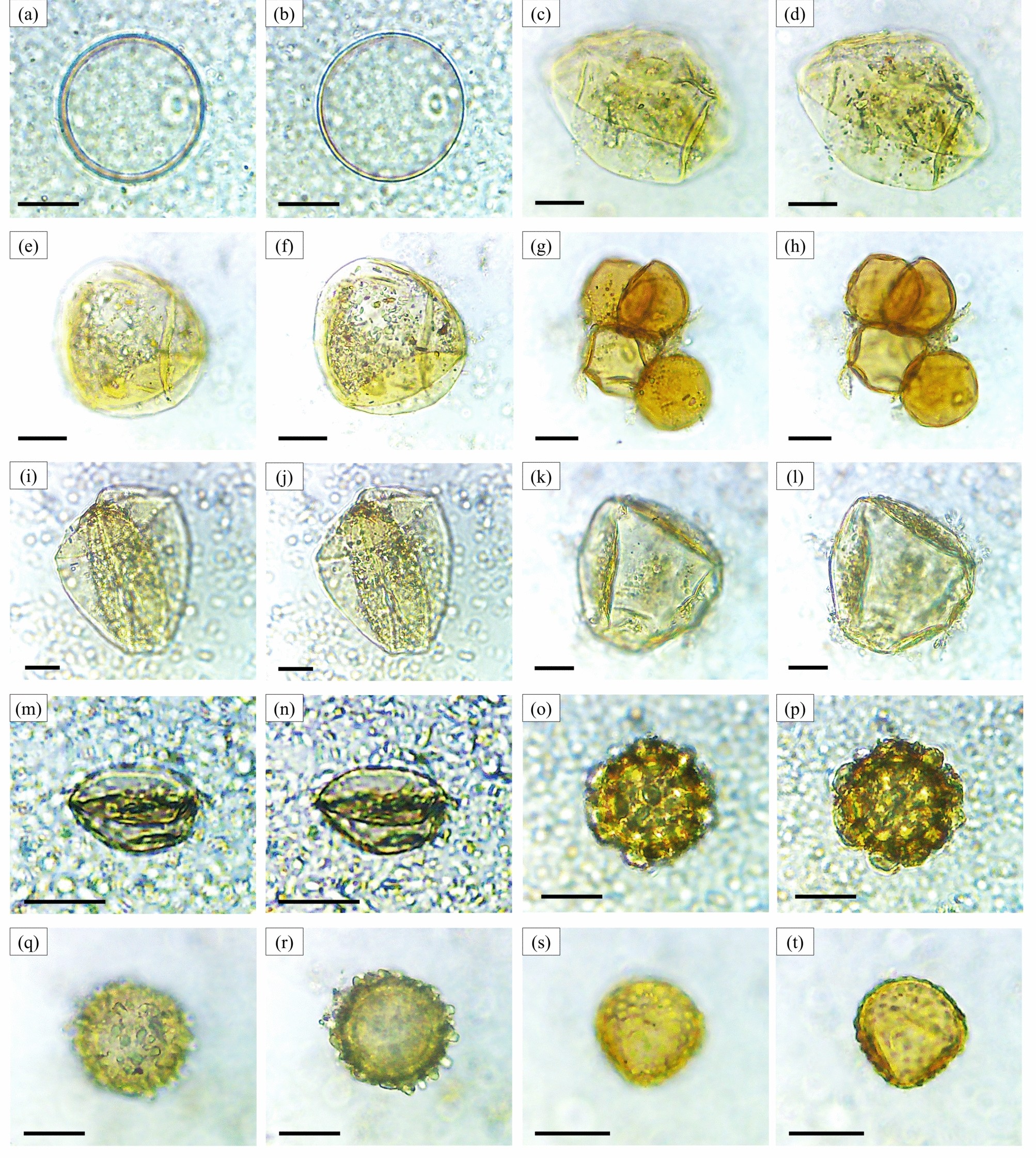
تصاویر میکروسکوپی از دانههای گرده (بزرگنمایی ۴۰۰ ×). بازدانگان: کاج ( a ، b )، سرو ( c ، d ). نهاندانگان: اولموس ( e ، f )، اولموس ( g ، h ) ، تاکستان ( i ، j )، ترم ( k ، l )، توسکا ( m ، n )، توس ( o ، p )، بلوط ( q ، r )، راش (fagaceae) ( s، t ) . مقیاس = ۱۰ میکرومتر. تمایزات مورفوتیپ از معیارهای شرح داده شده در شکل ۳ و تکمیلی پیروی میکنند.
تصاویر میکروسکوپی از دانههای گرده (بزرگنمایی ۴۰۰ ×). نهاندانگان: Fagaceae gen. indet. sp.2 ( a , b )، Aleurites sp.1 ( c , d )، Aleurites sp.2 ( e , f )، Acalypha sp. ( g , h )، Croton sp. ( i , j )، Euphorbiaceae gen. indet. ( k , l )، Lagerstroemia sp. ( m , n )، Schleichera sp. ( o , p )، Diospyros sp. ( q , r )، Oroxylum sp. ( s , t ). مقیاس = ۱۰ میکرومتر. تمایزات مورفوتیپ از معیارهای شرح داده شده در شکل ۳ و تکمیلی پیروی میکنند.

عکس میکروگراف از دانه های گرده (بزرگنمایی ۴۰۰ ×). آنژیوسپرم ها: Tilia sp. ( a , b ), Malvaceae gen. indet. ( ج ، د )، Spondias sp. ( e ، f )، Gynochthodes sp. ( g , h ), Arecaceae gen. indet. ( i , j ), Amaranthus sp. ( k , l ), Amaranthaceae gen. indet. sp.1 ( m ، n )، Amaranthaceae gen. indet. sp.2 ( o , p ), Amaranthaceae gen. indet. sp.3 ( q , r ), Zea mays ( s , t ). نوار مقیاس = ۱۰ میکرومتر. تمایز مورفوتایپ از معیارهای شرح داده شده در شکل ۳ و مکمل پیروی می کند.
تصاویر میکروسکوپی از دانههای گرده (بزرگنمایی ۴۰۰ ×). نهاندانگان: Poaceae gen. indet. sp.1 ( a ، b )، Poaceae gen. indet. sp.2 ( c ، d )، Poaceae gen. indet. sp.3 ( e ، f )، Poaceae gen. indet. sp.4 ( g ، h )، Poaceae gen. indet. sp.5 ( i ، j )، Poaceae gen. indet. sp.6 ( k ، l )، Physalis sp. ( m ، n )، Xanthium sp. ( o ، p )، Chromolaena sp. ( q ، r )، Carduus sp. ( s ، t ). مقیاس = ۱۰ میکرومتر. تمایزات مورفوتیپ از معیارهای شرح داده شده در شکل ۳ و تکمیلی پیروی میکنند.

تصاویر میکروسکوپی از دانههای گرده (بزرگنمایی ۴۰۰ ×). نهاندانگان: Asteraceae gen. indet. sp.1 ( a ، b )، Asteraceae gen. indet. sp.2 ( c ، d )، Asteraceae gen. indet. sp.3 ( e ، f )، Asteraceae gen. indet. sp.4 ( g ، h )، Asteraceae gen. indet. sp.5 ( i ، j )، Asteraceae gen. indet. sp.6 ( k ، l )، Asteraceae gen. indet. sp.7 ( m ، n )، Asteraceae gen. indet. sp.8 ( o ، p )، Amorphophallus sp. ( q ، r )، Araceae gen. indet. ( s ، t ). نوار مقیاس = ۱۰ میکرومتر. تمایزات مورفوتیپ از معیارهای شرح داده شده در شکل ۳ و تکمیلی پیروی میکنند.
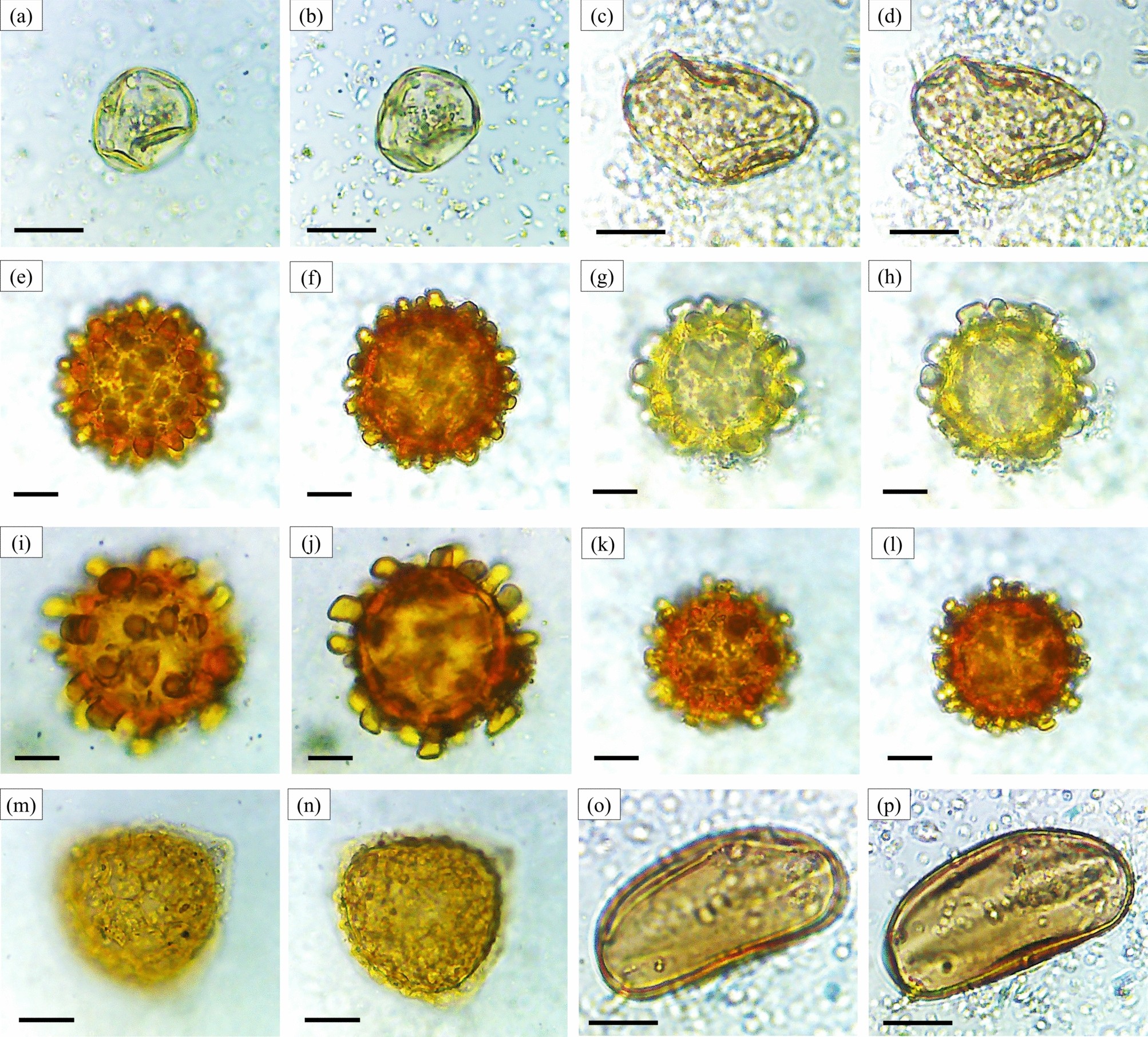
تصاویر میکروسکوپی از دانههای گرده (بزرگنمایی ۴۰۰ ×). نهاندانگان: Cyperus sp. ( a ، b )، Cyperaceae gen. indet. ( c ، d )، Hibiscus sp.1 ( e ، f )، Hibiscus sp.2 ( g ، h )، Hibiscus sp.3 ( i ، j )، Hibiscus sp.4 ( k ، l )، Vigna sp. ( m ، n )، Liliaceae gen. indet. ( o ، p ). نوار مقیاس = ۱۰ میکرومتر. تمایزات مورفوتیپ از معیارهای شرح داده شده در شکل ۳ و تکمیلی پیروی میکنند .
گرده درختی (AP)
گردههای درختی به طور مداوم در سراسر پروفیل نمونهبرداری شده نادر هستند و به صورت پراکنده و با فراوانی نسبی کم یافت میشوند. گونههای شناسایی شده شامل Schleichera sp. (Sapindaceae)، Diospyros sp. (Ebenaceae) و Spondias sp. (Anacardiaceae) هستند که به ترتیب در نمونههای جدا شده در اعماق ۰، ۳ و ۱۲ سانتیمتری یافت میشوند. Alnus sp. (Betulaceae) در عمق ۳ سانتیمتری شناسایی شده است. این سوابق، وقوع و توزیع عمودی انواع گردههای درختی را در این مجموعه مستند میکنند.
گرده غیر درختی (NAP)
گردههای غیردرختی بخش قابل توجهی از مجموعه گردهها را در تمام اعماق نمونهبرداری شده تشکیل میدهند. گونههای ثبتشدهی مکرر شامل Poaceae، Amaranthaceae، Asteraceae، Convolvulaceae، Cyperaceae و سایر گونههای علفی هستند. Zea mays (Poaceae) در عمق ۳ سانتیمتری وجود دارد و در طول تحقیقات میدانی نیز مشاهده شد. این گونهها نشاندهندهی وجود انواع گردههای علفی و زراعی در مواد نمونهبرداری شده هستند.
هاگها
هاگها در سراسر این پهنه به وفور یافت میشوند. گونههای ثبتشدهی رایج شامل گونههای Pteris (از تیرهی Pteridaceae)، Adiantum (از تیرهی Pteridaceae)، Dryopteris (از تیرهی Dryopteridaceae) و Lycopodium (از تیرهی Lycopodiaceae) هستند. این انواع هاگ، بخش عمدهای از مجموعهی پالینومورفها را در تمام اعماق تشکیل میدهند.
بحث
سیگنالهای پوشش گیاهی در چشماندازی که تحت تأثیر انسان قرار دارد
یکی از ویژگیهای برجسته مجموعه پالینومورفها، تسلط گونههای تولیدکننده اسپور، به ویژه مونیلوفیتها است که تقریباً ۴۳٪ از کل تعداد پالینومورفها را تشکیل میدهند. افزایش تعداد اسپورها معمولاً در خاکهای گرمسیری و نیمهگرمسیری و مجموعههای نزدیک به سطح گزارش میشود، جایی که سرخسها و لیکوفیتها اغلب به دلیل تولید زیاد اسپور، ساختارهای دیوارهای قوی و حفظ مطلوب نسبت به بسیاری از انواع گرده نهاندانگان، به طور نامتناسبی مشارکت دارند. ۳ ، ۲۷٫ فراوانی مورفوتیپهای مربوط به گونههای Pteris ، Blechnum ، Adiantum و Cyathea با توزیع گسترده آنها در زیستگاههای مرطوب زیرین و حاشیههای جنگلی در سراسر سرزمین اصلی آسیای جنوب شرقی سازگار است.
گرده گیاهان نهاندانه درختی نسبتاً کمتر از حد معمول است و تقریباً ۱۲٫۵٪ از مجموعه را تشکیل میدهد. الگوهای مشابهی در مطالعات مدرن باران گرده و خاک از مناظر گرمسیری اصلاحشده توسط انسان ثبت شده است، جایی که گردههای درختی اغلب توسط هاگهای فراوان محلی و گونههای غیر درختی رقیق میشوند ۲۸ ، ۲۹٫ در مجموعه حاضر، انواع گردههای درختی مانند Schleichera sp.، Diospyros sp.، Spondias sp.، Alnus sp. و اعضای خانوادههای Fagaceae و Ulmaceae به صورت پراکنده و با فراوانی نسبی کم یافت میشوند که حضور آنها را در منطقه منبع گرده بدون دلالت بر نمایش متناسب پوشش گیاهی ثابت، مستند میکند.
گردههای غیر درختی بخش قابل توجهی از این مجموعه را تشکیل میدهند، از جمله Poaceae، Amaranthaceae، Asteraceae، Convolvulaceae، Cyperaceae و سایر گونههای علفی. وجود گرده Zea mays ، همراه با گونههای کشتشده و رودرال مشاهدهشده در طول تحقیقات میدانی، سهم پوشش گیاهی کشاورزی و زمینهای باز را در مجموعه پالینومورفها مستند میکند. همانطور که به طور گسترده شناخته شده است، مجموعههای گرده خاک و نزدیک به سطح در مناظر آشفته، ورودیها را از لکههای گیاهی متعدد و مناطق منبع ادغام میکنند و بنابراین منعکسکننده ناهمگونی ترکیبی هستند تا حالتهای گسسته پوشش گیاهی ۱۶ ، ۳۰ .
نکته مهم این است که تغییرپذیری عمودی در پروفیل خاک ۳۰ سانتیمتری نباید به عنوان یک توالی زمانی منظم تفسیر شود. در غیاب دادههای مستقل کنترل سن یا نرخ رسوبگذاری، عمق در پروفیلهای خاک کمعمق مستقیماً با زمان مطابقت ندارد. در عوض، تفاوتهای مشاهدهشده از نظر عمق، نشاندهنده تنوع چینهشناسی در یک مجموعه یکپارچهشده با زمان و فیلترشده از نظر تافونومیکی است که تحت تأثیر آشفتگی زیستی، فعالیت ریشه و بازسازیهای دورهای قرار دارد. در نتیجه، این مجموعه در اینجا به عنوان مستندسازی دامنه و همزیستی انواع پالینومورفهای موجود در یک موزاییک جنگل-کشاورزی تحت تأثیر انسان تفسیر میشود، نه گذارهای اکولوژیکی، مراحل جانشینی یا تغییر کاربری زمین تاریخی.
روی هم رفته، وقوع همزمان گردههای درختی، گردههای غیر درختی و هاگهای فراوان، ماهیت ناهمگن ورودی گرده را در مناظر کوهستانی آشفته برجسته میکند. این دادهها به جای بازسازی پویایی پوشش گیاهی در طول زمان، اطلاعات زمینهای در مورد نمایش پالینومورفها در خاکهای یک محیط مختلط جنگلی-کشاورزی ارائه میدهند و ارزش مجموعه دادههای مرجع مبتنی بر مورفولوژی را برای تفسیر محیطهای رسوبی قابل مقایسه در مناطق گرمسیری تقویت میکنند. توصیفات مربوط به کاربری اراضی امروزی صرفاً به عنوان پسزمینه زمینهای ارائه شده و برای کالیبراسیون یا اعتبارسنجی مجموعه گردهشناسی استفاده نمیشوند، در نتیجه از استنتاج دایرهای بین مشاهده پوشش گیاهی و تفسیر گرده جلوگیری میشود.
سهم پالینولوژیکی در دادههای مرجع منطقهای
مطالعه حاضر یکی از جامعترین توصیفات ریختشناسی گرده و هاگ را از یک جنگل برگریز مخلوط تخریبشده در شمال تایلند ارائه میدهد. این تحقیق با مستندسازی ۳۷ خانواده گیاهی در میان لیکوفیتها، بریوفیتها، مونیلوفیتها، بازدانگان و نهاندانگان، چارچوب مرجع پالینولوژیکی منطقهای را برای اکوسیستمهای کوهستانی سرزمین اصلی جنوب شرقی آسیا به طور قابل توجهی گسترش میدهد. استفاده از استخراج غیر استولیز و نمونهبرداری با وضوح خوب، بازیابی و شناسایی مجموعهای غنی از نظر طبقهبندی و از نظر ریختشناسی متنوع از پالینومورفها را امکانپذیر کرده است که بسیاری از آنها در پایگاههای داده جهانی موجود کمتر نشان داده شدهاند.
این مطالعه چندین کمک قابل توجه به تحقیقات پالینولوژی منطقهای ارائه میدهد. این مطالعه دادههای ریختشناسی با وضوح بالا را در سطوح جنس و خانواده از یک چشمانداز دستخورده توسط انسان ارائه میدهد – نوعی منطقه که در مقایسه با جنگلهای دستنخورده یا توربزارهای دستنخورده در مطالعات قبلی کمتر مورد توجه قرار گرفته است. علاوه بر این، مجموعه دادهها شامل طیف گستردهای از گونهها، از جمله جنسهای مرتبط با اکولوژی و متنوع از نظر ریختشناسی مانند Gynochthodes sp.، Aleurites spp.، Physalis sp. و انواع مختلف Asteraceae ناشناخته است. این گونهها اغلب در بررسیهای منطقهای موجود کمتر مورد توجه قرار میگیرند یا فقط به طور خلاصه ذکر میشوند، که نشاندهنده ارزش این مجموعه دادهها برای افزایش ارزیابیهای تنوع زیستی و اطلاعرسانی در تحقیقات مرتبط با حفاظت است.
محدودیتهای زمانی و زمانی پالینولوژی خاکهای کمعمق
از آنجا که مواد مورد تجزیه و تحلیل از یک پروفیل خاک کمعمق به جای یک توالی دریاچهای یا پیت که به طور مداوم در حال تجمع است، میآید، این مجموعه باید به عنوان یک سیگنال یکپارچه زمانی و فیلتر شده از نظر تافونومیکی از ورودیهای محلی-فرامحلی تفسیر شود. تخریب افتراقی میتواند به طور انتخابی دانههای دیواره نازک یا با تزئینات ضعیف را حذف کند، در حالی که گونههای قوی ممکن است بیش از حد نمایان شوند و اختلاط فیزیکی (به عنوان مثال، زیستتخریب و فعالیت ریشه) میتواند تضادهای چینهشناسی را محو کند. شواهد تجربی نشان میدهد که حفظ در رسوبات حرا و گرمسیری به شدت وابسته به گونه است ۳۱ ، و سنتزهای گستردهتر نشان میدهند که دیاژنز و پردازش آزمایشگاهی میتوانند در تعامل باشند تا آنچه را که در نهایت حفظ و مشاهده میشود، شکل دهند ۳۲٫ فرآیندهای مدرن انتقال و رسوبگذاری در محیطهای مرطوب گرمسیری میتوانند بر منطقه منبع و نمایش تأثیر بیشتری بگذارند و ارزش بررسیهای پوشش گیاهی جفت شده و نمونهبرداری تکراری را هنگام تفسیر مجموعههای خاک و نزدیک به سطح تقویت کنند ۳۰ . بر این اساس، مقایسه مستقیم با سوابق گرده منطقهای حاصل از دریاچهها، زمینهای باتلاقی یا هستههای رسوبی با قدمت چینهشناسی باید با احتیاط انجام شود، زیرا چنین بایگانیهایی اساساً از نظر تداوم رسوبگذاری، وضوح زمانی و حساسیت به اختلاط پس از رسوبگذاری با پروفیلهای خاک متفاوت هستند.
این فرآیندهای تافونومیک در کنار هم، قابلیت تفسیر الگوهای فراوانی نسبی را محدود میکنند و این نکته را تقویت میکنند که مجموعههای پالینومورف خاک در درجه اول منعکسکننده حضور ترکیبی هستند تا ساختار کمی پوشش گیاهی.
ملاحظات روششناختی و فناوری
یکی از نقاط قوت عملی رویکرد مورفوتیپ-کاتالوگ این است که میتواند از شناسایی تکرارپذیر پشتیبانی کند و در عین حال مواردی را که تخصیصهای سطح گونه به طور مداوم در بین تحلیلگران، میکروسکوپها و فلورهای منطقهای قابل دستیابی نیستند، در بر بگیرد. تعاریف هماهنگ مورفوتیپ، مترادفهای واضح و ابردادههای مرتبط با تصویر به طور فزایندهای به عنوان پیشنیازهای ترکیب مجموعه دادههای گردهشناسی در سایتها و مطالعات ۳۳ ، ۳۴ شناخته میشوند . در زمینههای هوازی-زیستی و پالئواکولوژیکی، مهارت تحلیلگر و کنترل کیفیت شفاف بر سازگاری در سطح تاکسون تأثیر میگذارند ۳۵ ، ۳۶٫ بنابراین، این مطالعه توصیه میکند که مجموعه دادههای مورفوتیپ (تکمیلی) به عنوان یک منبع مرجع نسخهبندی شده (با شناسههای پایدار، معیارهای تشخیصی و میکروگرافهای نماینده) در نظر گرفته شود تا بهروزرسانیهای آینده، مقایسه بین آزمایشگاهها و ادغام در پایگاههای داده گرده منطقهای تسهیل شود.
فراتر از بهبود دید مورفولوژیکی، انتخاب روش تصفیه آزمایشگاهی پیامدهای گستردهتری برای تجزیه و تحلیلهای پاییندستی و مقایسهپذیری دارد. کارهای تجربی اخیر نشان میدهد که مراحل شیمیایی رایج – بهویژه استولیز – میتوانند ترکیب مولکولی اسپورومورفها را به طور قابل توجهی تغییر دهند و ممکن است گونههای مختلف را به طور غیر یکنواخت تحت تأثیر قرار دهند ۳۷ ، در حالی که دیاژنز و پردازش میتوانند با هم تعامل داشته باشند تا آنچه را که در نهایت در زیر میکروسکوپ حفظ و مشاهده میشود، شکل دهند ۳۲٫ در جایی که هدف اصلی مستندسازی تزئینات ظریف برای توسعه اطلس و آموزش مبتنی بر تصویر است، به حداقل رساندن مراحل اکسیداسیون تهاجمی میتواند به حفظ ویژگیهای ظریفی که در غیر این صورت ممکن است تغییر یافته یا از بین بروند، کمک کند.
اتخاذ روش استخراج غیر استولیز (به دنبال ۲۴ ) یکی دیگر از مزایای روششناختی را نشان میدهد. برخلاف استولیز سنتی، که خطر تخریب ساختارهای ظریف اگزین – به ویژه در گردههای خاردار، مشبک یا وروکات – را به همراه دارد، این رویکرد جزئیات مورفولوژیکی دقیقی را که برای شناسایی دقیق طبقهبندی ضروری است، حفظ میکند. در نتیجه، عکسهای میکروسکوپی با کیفیت بالا و توصیفات مورفومتریک تولید شده در این مطالعه نه تنها دادههای طبقهبندی را ارائه میدهند، بلکه به عنوان یک اطلس مرجع برای تحقیقات آینده در پالینولوژی گرمسیری، چه آنالوگ و چه دیجیتال، نیز عمل میکنند.
اهمیت این کار همچنین در کاربرد بالقوه آن برای سیستمهای تشخیص خودکار گرده مشهود است. از آنجایی که فناوریهای یادگیری ماشین و بینایی کامپیوتر به طور فزایندهای در گردهشناسی به کار میروند، در دسترس بودن دادههای آموزشی متنوع و منطقهای، محدودیت اصلی در قابلیت انتقال مدل است ۳۸ ، ۳۹ ، ۴۰ ، ۴۱٫ با تکیه بر تلاشهای قبلی شناسایی خودکار ۴۲ ، ۴۳ ، ۴۴ ، مجموعه دادههای تصویری گسترده تولید شده در اینجا – مرتبط با تعاریف مورفوتیپ و زمینه اکولوژیکی – پایه و اساس ضروری برای توسعه و محک زدن الگوریتمها در مناطق گرمسیری و نیمهگرمسیری را فراهم میکند که در آنها کتابخانههای آموزشی دیجیتال پراکنده هستند.
به طور خلاصه، این مطالعه سهم قابل توجهی در حوزه پالینولوژی گرمسیری دارد. این مطالعه پایگاههای داده گرده منطقهای را بهبود میبخشد، یک مبنای مورفولوژیکی حیاتی برای مناظر جنگلی آشفته فراهم میکند و شکاف بین طبقهبندی سنتی و روشهای محاسباتی نوظهور را پر میکند. فراتر از ارتباط مستقیم آن با منطقه مورد مطالعه، دقت روششناختی و عمق اکولوژیکی این تحقیق، آن را به طور گسترده برای سایر مناطق کوهستانی و تغییر یافته توسط انسان در جنوب شرقی آسیا و فراتر از آن قابل اجرا میکند. با تمرکز بر مورفوتیپهای قابل انتقال و تصاویر قابل استفاده مجدد، این کار به گونهای طراحی شده است که با تقویت مبنای شناسایی و امکان مقایسه متقابل مطالعات، بازسازیهای چینهشناسی طولانیتر از منطقه را تکمیل کند – نه اینکه آنها را تکرار کند.
نتیجهگیری
این مطالعه، مستندسازی ریختشناسی-محور انواع گرده و هاگ بازیابیشده از یک پروفیل خاک کمعمق در یک موزاییک مخلوط جنگل-کشاورزیِ متأثر از فعالیتهای انسانی در شمال تایلند را ارائه میدهد. با استفاده از یک پروتکل استخراج غیر استولیز و نمونهبرداریهای جزئی با وضوح بالا، گرده و هاگهای نماینده ۳۷ خانواده گیاهی شناسایی شدند که شامل لیکوفیتها، بریوفیتها، مونیلوفیتها، بازدانگان و نهاندانگان میشوند. گونههای تولیدکننده هاگ در این مجموعه غالب هستند، در حالی که انواع گرده نهاندانگان درختی و غیر درختی در فراوانی نسبی کمتری یافت میشوند.
از آنجا که مواد مورد تجزیه و تحلیل از یک پروفیل خاک کمعمق و بدون کنترل زمانی مستقل مشتق شدهاند، این مجموعه باید به عنوان یک سیگنال یکپارچه زمانی و فیلتر شده از نظر تافونومیکی تفسیر شود، نه به عنوان سابقهای از تغییر پوشش گیاهی در طول زمان. بر این اساس، سهم اصلی این مطالعه در نقش آن به عنوان یک مجموعه داده مرجع مورفولوژیکی منطقهای است که توسط توصیفات مورفوتیپ استاندارد، عکسهای میکروسکوپی با کیفیت بالا و مستندات طبقهبندی شفاف پشتیبانی میشود.
با گسترش منابع مرجع موجود برای گرده و هاگ از موزاییکهای جنگل-کشاورزیِ تحت تأثیر انسان، این کار، اطلسهای مورفولوژی موجود و مطالعات مدرن گرده را که عمدتاً بر محیطهای دستنخورده یا چینهشناسیِ مشخص تمرکز دارند، تکمیل میکند. هنگامی که این دادهها با سوابق رسوبیِ تاریخگذاریشدهی مستقل، طرحهای نمونهبرداریِ تکرارشونده یا بررسیهای پوشش گیاهیِ جفتشده ترکیب شوند، میتوانند از تجزیه و تحلیلهای دیرینهمحیطشناسی، فلوریستیک و مقایسهایِ قویتری پشتیبانی کنند. آرشیو تصاویرِ گردآوریشده همچنین پایهای برای آموزش و محکزنیِ رویکردهای خودکارِ تشخیص گرده در مناطق گرمسیری فراهم میکند که در آنها کتابخانههای مرجع دیجیتال محدود هستند.
در دسترس بودن دادهها
دادههای پشتیبان یافتههای این مطالعه، از جمله مجموعه دادههای مورفوتایپ (تکمیلی)، در اطلاعات تکمیلی ارائه شدهاند. مطالب اضافی در صورت درخواست منطقی از نویسنده مسئول در دسترس هستند.
منابع
-
دای، ال. و همکاران. ریختشناسی گرده گیاهان گرمسیری منتخب در شبهجزیره مالزی و تأثیر آن در بازسازی دیرینهبومشناسی جنوب شرقی آسیا. Rev. Palaeobot. Palynol. ۳۱۶ ، ۱۰۴۹۳۵٫ https://doi.org/10.1016/j.revpalbo.2023.104935 (۲۰۲۳).
-
روهان، جی. و گادئول، ام. طبقهبندی گیاهان: یک دیدگاه تاریخی، چالشهای فعلی و چشماندازها. در طبقهبندی مولکولی گیاهان (ویراستار بس، پی.) (هومانا، ۲۰۲۱).
-
هالبریتر، اچ. و همکاران. اصطلاحات مصور گردهشناسی، ویرایش دوم (اشپرینگر، ۲۰۱۸).
-
پانت، دبلیو. و همکاران. واژهنامه اصطلاحات گرده و هاگ. Rev. Palaeobot. Palynol. ۱۴۳ ، ۱–۸۱ (۲۰۰۷).
-
چنگ، ز. و همکاران. مطالعهای بر روی باران گرده مدرن و مورفولوژی گرده در شبهجزیره مالای غربی گرمسیری و پیامدهای آن برای بازسازیهای دیرینهمحیطی در منطقه سوندا. Rev. Palaeobot. Palynol. ۲۷۹ ، ۱۰۴۲۳۶٫ https://doi.org/10.1016/j.revpalbo.2020.104236 (۲۰۲۰).
-
تیره، پ. گرده و بقایای گیاهی ماکروسکوپی به عنوان شاخصهای محیطهای محلی و منطقهای. در کتابچه راهنمای علوم باستانشناسی، ویرایش دوم (ویراستاران پولارد، ای. ام. و همکاران) ۱–۱۸ (وایلی، ۲۰۲۳).
-
لدرو، ام. و همکاران. تخریب تخمینی Caatinga بر اساس باران گرده مدرن نهشته شده در مخازن. ربات Acta. سوتین. ۳۶ , e20220111. https://doi.org/10.1590/1677-941x-abb-2022-0111 (۲۰۲۲).
-
هوفکووا، پ. و همکاران. رویدادهای اقلیمی هولوسن مرتبط با تغییرات محیطی در حوضه دریاچه کوموری، جمهوری چک. هولوسن ۲۷ ، ۱۱۳۲-۱۱۴۵ (۲۰۱۷).
-
بوش، امبی. در تفسیر گردههای فسیلی گندمیان در مناطق پست و مرطوب نئوتروپیک. پالئوژئوگر. پالئوکلیماتول. پالئوکول. ۱۷۷ ، ۵-۱۷ (۲۰۰۲).
-
بیرک، اچ اچ و بیرک، اچ جی بی، پالئواکولوژی کواترنری (ادوارد آرنولد، ۱۹۸۰).
-
هارا، م. و همکاران. ترکیب فلوریستیک جنگلهای کوهستانی گرمسیری در پارک ملی اینتانون، شمال تایلند. Nat. Hist. Res. ۷ ، ۱–۱۷ (۲۰۰۲).
-
گاردنر، س.، سیدیسانتورن، پ. و آنوسارنسانتورن، و. راهنمای میدانی درختان جنگلی شمال تایلند (پروژه انتشارات کوبفای، ۲۰۰۰).
-
ماکسول، جی. اف. پوشش گیاهی و فلور عروقی پارک ملی سوتپ-پوی، شمال تایلند. مجله تایلندی. زیستغارگران. ۵ ، ۱-۲۰۵ (۲۰۰۴).
-
ساترابوروت، تی. و همکاران. کاهش سریع ذخایر کربن آلی خاک پس از تبدیل جنگل به مزرعه ذرت در یک حوزه آبخیز در شمال تایلند. Environ. Challenges ۱۷ ، ۱۰۱۰۴۲٫ https://doi.org/10.1016/j.envc.2024.101042 (۲۰۲۴).
-
کرواسیلیپ، جی. و همکاران. ارزیابی تغییرات کاربری زمین و پوشش زمین در استان نان، تایلند، با استفاده از دادههای ماهوارهای چند حسگری و موتور گوگل ارث. محیط زیست. منابع طبیعی. مجله ۲۱ ، ۱۸۶-۱۹۷ (۲۰۲۳).
-
مرکوری، ای. ام. و همکاران. شاخصهای گرده انسانی از مکانهای باستانشناسی به عنوان شواهد محلی از محیطهای ناشی از فعالیت انسان در شبه جزیره ایتالیا. مجله Ann. Bot. ۳ ، ۱۴۳–۱۵۳ (۲۰۱۳).
-
ساترابوروت، تی. و همکاران. بازسازی تغییر جنگل در ارتفاعات تایلند: شواهدی از اختلالات انسانی، کشاورزی و احیای اکولوژیکی. درخت برای مردم ۲۲ ، ۱۰۱۰۰۰٫ https://doi.org/10.1016/j.tfp.2025.101000 (۲۰۲۵).
-
اداره پارکهای ملی، حیات وحش و حفاظت از گیاهان. پارک ملی سری نان. http://nps.dnp.go.th/parksdetail.php?id=99 (۲۰۲۳).
-
اداره جنگلداری سلطنتی، تایلند. دادههای اطلاعات جنگل – آمار جنگل، BE 2543 (2000). https://forestinfo.forest.go.th/Content.aspx?id=158 (۲۰۲۵).
-
انجمن QGIS. سیستم اطلاعات جغرافیایی QGIS (نسخه ۳٫۲۲٫۱۰). https://qgis.org (۲۰۲۲).
-
دفتر هماهنگی امور بشردوستانه سازمان ملل متحد. تایلند – مرزهای اداری فراملی. https://data.humdata.org/dataset/cod-ab-tha (۲۰۲۳).
-
وزارت منابع آب. مجموعه دادههای نقشههای ملی. https://opendata.onde.go.th/en/dataset/9-national-maps (۲۰۲۲).
-
Copernicus Sentinel-2 (پردازش شده توسط ESA). محصول بازتاب BOA سطح ۲A MSI. https://doi.org/10.5270/S2_-znk9xsj (۲۰۲۱).
-
دی آلمیدا سانتوس، آر. و لدرو، ام پی، پروتکل بدون اسید برای استخراج گرده از رسوبات کواترنر. پالینولوژی ۴۶ ، ۱۹۶۰۹۱۶ (۲۰۲۲).
-
بوچال، جی. ام. و همکاران. مطالعه کیفی LM و SEM از پالینوفلورای Messel: بخش اول. گرانا ۶۳ ، ۱۹۳-۲۴۶ (۲۰۲۴).
-
بوچال، جی. ام. و همکاران. مطالعه کیفی LM و SEM از پالینوفلورای Messel: بخش دوم. Rev. Palaeobot. Palynol. ۳۱۹ ، ۱۰۵۳۴۹٫ https://doi.org/10.1016/j.revpalbo.2025.105349 (۲۰۲۵).
-
Tryon، AF & Lugardon، BL Spores of Pteridophyta (اسپرینگر، ۱۹۹۱).
-
ژنگ، ز. و همکاران. تأثیرات انسانی بر تغییر پوشش زمین در اواخر هولوسن و از بین رفتن تنوع زیستی فلوریستیک در مناطق گرمسیری جنوب شرقی آسیا. مجموعه مقالات Natl Acad. Sci. USA ۱۱۸ ، e2022210118. https://doi.org/10.1073/pnas.2022210118 (۲۰۲۱).
-
فنگ، جی. و همکاران. اختلالات انسانی، ساختار جامعه درختی فیلوژنتیکی و عملکردی را شکل میدهد. برای. مدیریت محیط زیست. ۳۱۳ ، ۱۸۸-۱۹۸٫ https://doi.org/10.1016/j.foreco.2013.10.047 (۲۰۱۳).
-
ماس، پی تی، کرشاو، ای پی و گریندرود، جی اف. انتقال و رسوب گرده در محیطهای رودخانهای و دریایی در مناطق گرمسیری مرطوب شمال شرقی استرالیا. Rev. Palaeobot. Palynol. ۱۳۴ ، ۵۵-۶۹ (۲۰۰۵).
-
Phuphumirat، W. et al. تخریب گرده در رسوبات حرا. کشیش Palaeobot. پالینول. ۲۲۱ ، ۱۰۶-۱۱۶٫ https://doi.org/10.1016/j.revpalbo.2015.06.004 (۲۰۱۵).
-
زاوادا، اماس و هکلی، پیسی. تأثیر دیاژنز و استولیز بر حفظ ریختشناسی گرده. Rev. Palaeobot. Palynol. ۳۰۲ ، ۱۰۴۶۷۹٫ https://doi.org/10.1016/j.revpalbo.2022.104679 (۲۰۲۲).
-
بیرکس، اچجیبی و همکاران. رویکردهایی برای هماهنگسازی طبقهبندی گرده در پالینولوژی کواترنری. Rev. Palaeobot. Palynol. ۳۱۹ ، ۱۰۴۹۸۹٫ https://doi.org/10.1016/j.revpalbo.2023.104989 (۲۰۲۳).
-
فلانتوا، SGA و همکاران. راهنمایی برای پردازش و استانداردسازی دادههای پالئواکولوژیکی جهانی. Glob. Ecol. Biogeogr. ۳۲ ، ۱۳۷۷-۱۳۹۴٫ https://doi.org/10.1111/geb.13693 (۲۰۲۳).
-
گالن، سی. و همکاران. پایش گرده: حداقل الزامات و تکرارپذیری تجزیه و تحلیل. Aerobiologia ۳۰ ، ۳۸۵-۳۹۵ (۲۰۱۴).
-
سیکوپاریجا، ب. و همکاران. پایش گرده: آزمایش مهارت تحلیلگر. Aerobiologia ۳۳ ، ۱۹۱–۱۹۹ (۲۰۱۷).
-
خفاشها، YF و همکاران. تأثیر عملیات شیمیایی بر ترکیب اسپورومورف. علوم زیستی ۲۲ ، ۴۶۸۹-۴۷۰۴٫ https://doi.org/10.5194/bg-22-4689-2025 (۲۰۲۵).
-
هولت، کی.ای. و بنت، کی.دی. اصول و روشهای گردهشناسی خودکار. نیو فیتول. ۲۰۳ ، ۷۳۵-۷۴۲٫ https://doi.org/10.1111/nph.12848 (۲۰۱۴).
-
بارنز، سیام و همکاران. طبقهبندی گرده خودکار قیاسی با استفاده از یادگیری عمیق. نیو فیتول. ۲۴۰ ، ۱۳۰۵-۱۳۲۶٫ https://doi.org/10.1111/nph.19186 (۲۰۲۳).
-
دوراند، ام. و همکاران. شناسایی گرده از طریق شبکههای عصبی کانولوشن. PLoS ONE ۱۹ ، e0302424. https://doi.org/10.1371/journal.pone.0302424 (۲۰۲۴).
-
فنگ، جی تی و همکاران. تشخیص و جداسازی گرده در اسلایدهای پالینولوژیکی با یادگیری عمیق. پالئوبیولوژی ۵۱ (۳۹۴)، ۴۰۷٫ https://doi.org/10.1017/pab.2025.10059 (۲۰۲۵).
-
پونیاسنا، اس. دبلیو. و همکاران. شناسایی خودکار نمونههای گرده متنوع نئوتروپیکال. روشها Ecol. Evol. ۱۳ ، ۲۰۴۹–۲۰۶۴٫ https://doi.org/10.1111/2041-210X.13917 (۲۰۲۲).
-
خانژینا، ن. و همکاران. تشخیص دانه گرده با استفاده از شبکههای عصبی کانولوشن. ESANN Proc. ۴۰۹–۴۱۴ (۲۰۱۸).
-
رستمی، ام.ای و همکاران. یادگیری عمیق برای طبقهبندی دقیق دانههای گرده مخروطیان. فرانت. بیگ دیتا ۸ ، ۱۵۰۷۰۳۶. https://doi.org/10.3389/fdata.2025.1507036 (۲۰۲۵).
تقدیرنامهها
این تحقیق از طریق کمک مالی شورای ملی تحقیقات تایلند (NRCT) تحت صندوق تحقیقات و نوآوری برای سال مالی ۲۰۲۴، از طریق کمک هزینه توسعه محققان جوان (شماره کمک هزینه N42A670907) پشتیبانی مالی شده است. همچنین از پارک و اداره پارکهای ملی، حیات وحش و حفاظت از گیاهان برای تسهیل دسترسی به منطقه مورد مطالعه و ارائه کمکهای ارزشمند در طول جمعآوری دادههای میدانی، صمیمانه قدردانی میشود. تشکر ویژه از آزمایشگاه دانشکده مطالعات محیط زیست و منابع دانشگاه ماهیدول برای ارائه امکانات آزمایشگاهی، تجهیزات و همکاری علمی در تجزیه و تحلیل گردهشناسی، و همچنین از کادر دانشگاهی برای راهنماییهای ارزشمند و مشاوره فنی آنها. همچنین از ویراستار و داوران ناشناس برای نظرات سازندهشان که به طور قابل توجهی وضوح و دقت علمی نسخه خطی را بهبود بخشید، سپاسگزاریم.
بودجه
این پروژه توسط شورای ملی تحقیقات تایلند (NRCT) (شماره کمک هزینه N42A670907) تأمین مالی شده است. بودجه دسترسی آزاد توسط دانشگاه ماهیدول فراهم و سازماندهی شده است.
اعلامیههای اخلاقی
منافع رقابتی
نویسندگان هیچ گونه تضاد منافعی را اعلام نمیکنند.
اطلاعات تکمیلی
یادداشت ناشر
اشپرینگر نیچر در مورد ادعاهای مربوط به صلاحیت قضایی در نقشههای منتشر شده و وابستگیهای سازمانی بیطرف باقی میماند.
اطلاعات تکمیلی
حقوق و مجوزها
دسترسی آزاد این مقاله تحت مجوز بینالمللی Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 منتشر شده است که هرگونه استفاده، اشتراکگذاری، توزیع و تکثیر غیرتجاری را در هر رسانه یا قالبی مجاز میداند، مادامی که به نویسنده(گان) اصلی و منبع، اعتبار کافی بدهید، پیوندی به مجوز Creative Commons ارائه دهید و مشخص کنید که آیا محتوای دارای مجوز را اصلاح کردهاید یا خیر. شما تحت این مجوز اجازه اشتراکگذاری محتوای اقتباسشده برگرفته از این مقاله یا بخشهایی از آن را ندارید. تصاویر یا سایر مطالب شخص ثالث در این مقاله در مجوز Creative Commons مقاله گنجانده شدهاند، مگر اینکه در خط اعتباری مطلب، طور دیگری ذکر شده باشد. اگر مطلبی در مجوز Creative Commons مقاله گنجانده نشده باشد و استفاده مورد نظر شما طبق مقررات قانونی مجاز نباشد یا از استفاده مجاز فراتر رود، باید مستقیماً از دارنده حق چاپ اجازه بگیرید. برای مشاهده نسخهای از این مجوز، به http://creativecommons.org/licenses/by-nc-nd/4.0/ مراجعه کنید .
درباره این مقاله

به این مقاله استناد کنید
ساترابوروت، ت.، وونگواسانا، س.، پوتای، ت. و همکاران. تنوع مورفولوژیکی گرده و هاگ در یک موزاییک جنگل-کشاورزی کوهستانی تحت تأثیر انسان در شمال تایلند. Sci Rep ۱۶ ، ۶۷۹۴ (۲۰۲۶). https://doi.org/10.1038/s41598-026-37899-9
- دریافت شده
- پذیرفته شده
- منتشر شده
- نسخه رکورد
- DOIhttps://doi.org/10.1038/s41598-026-37899-9
کلمات کلیدی
- پالینومورفها
- مجموعه دادههای مرجع پالینولوژیکی
- آماده سازی غیر استولیز
- موزاییک جنگلهای گرمسیری-کشاورزی
- آسیای جنوب شرقی